

Bt, Bc et Cd peuvent déclencher diverses voies de développement qui leur permettent de persister dans leur niches. En particulier, ces bactéries peuvent produire des biofilms, former des spores ou entrer dans un état de persistance non sporulé. Ces processus sont activés lorsque les bactéries rencontrent des conditions environnementales défavorables lors d’une infection ou à l’extérieur de l’hôte.
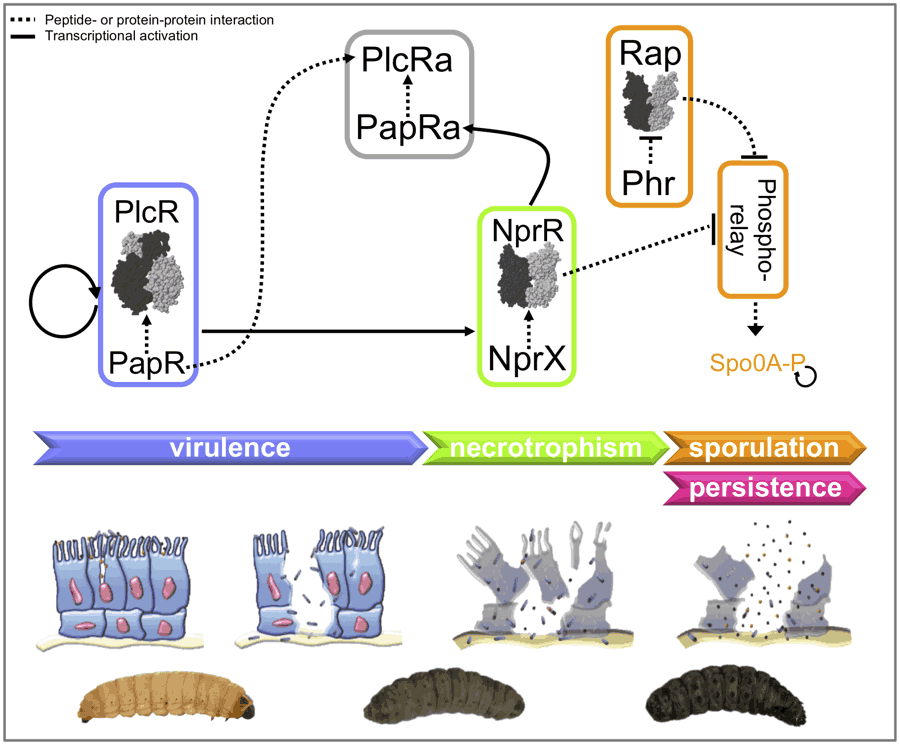
Les différentes étapes du cycle infectieux de Bt/Bc (virulence, nécrotrophisme et sporulation) sont contrôlées par la communication entre cellules ou les systèmes de communication du quorum-sensing (QS) qui permettent aux bactéries de coordonner l’expression génique en réponse à la densité cellulaire et aux stimuli environnementaux(doi: 10.3390/toxins6082239). Nos recherches ont conduit à la caractérisation de la famille de capteurs de quorum RRNPP, composée initialement de Rap, NprR, PlcR et PrgX(doi: 10.1073/pnas.0704501104). Des travaux sont actuellement en cours pour étudier comment le QS fonctionne in vivo au niveau de la cellule unique. Nous caractérisons également des molécules capables d’inhiber le QS (via l’extinction du quorum) et ainsi réduire la virulence bactérienne ou la sporulation en collaboration avec le Pr Z. Hayouka (Université hébraïque de Jérusalem). De plus, le rôle écologique des systèmes de QS sera étudié en utilisant des approches de biologie évolutive en collaboration avec le Pr. B. Raymond (Université d’Exeter).
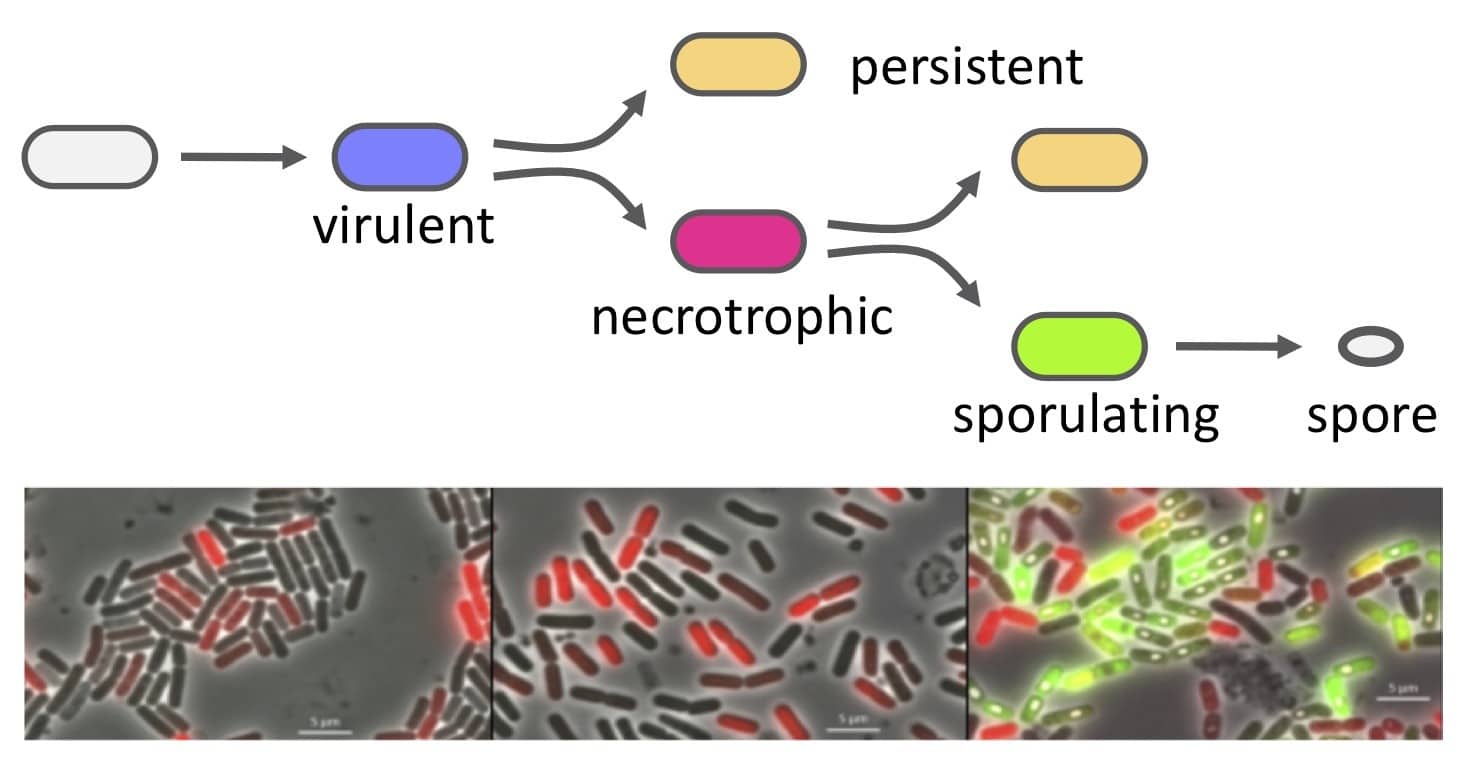

Les souches Bt canoniques produisent des cristaux insecticides parasporaux pendant la sporulation dans la même cellule que la spore. Quelques rares souches peuvent cependant se différencier en deux sous-populations distinctes de formateurs de spores et de producteurs de cristaux, et nous avons montré que ce phénotype de division du travail confère à la bactérie un avantage en termes de fitness dans la compétition avec une souche Bt canonique (doi : 10.1038/ismej.2014.122). Le régulateur de transcription CpcR a été caractérisé comme étant responsable de ce phénotype (doi : 10.1111/mmi.14439). Les travaux en cours visent à comprendre les mécanismes sous-jacents à cette hétérogénéité phénotypique. Ces travaux illustrent la diversité des stratégies employées par ces bactéries pour contrôler leur sporulation et permettre leur dissémination dans différentes niches écologiques. Ces études ont été réalisées en collaboration avec le laboratoire du Pr. F. Song (IPP CAAS Beijing) et ont contribué à la création du Laboratoire International Associé centré sur les interactions bactéries-plantes-insectes pour le biocontrôle des maladies (LIA BIPI).

Un composant majeur du biofilm est la matrice, une substance de type gel produite par les bactéries elles-mêmes et dans laquelle elles sont intégrées. Chez Bt, nous avons montré que la matrice du biofilm est composée de deux protéines (TasA et CalY) et de deux exopolysaccharides (Cps et Eps). En plus de leur rôle structurel dans la biosynthèse de la matrice, CalY et Cps sont également impliqués dans la pathogénicité de la bactérie. CalY peut fonctionner comme une adhésine, et Cps forme une capsule entourant la bactérie capable de se lier à des surfaces inertes ou vivantes. Eps et Cps sont distribués de manière très hétérogène dans le biofilm. Cps est localisé uniquement à la périphérie, en contact avec les surfaces solides, tandis qu’Eps est localisé uniquement au centre. Nous aimerions comprendre comment la transition vers une production uniquement de Cps ou Eps est régulée.
La persistance du biofilm (adhérence, protection contre les agents externes) et la résistance de la spore présente dans les biofilms font de Bc un problème sérieux dans l’industrie agroalimentaire. Nous nous intéressons particulièrement à déterminer l’influence des facteurs rencontrés dans les unités de production (milieu, pH, température, présence d’autres organismes) sur la sporulation des biofilms.
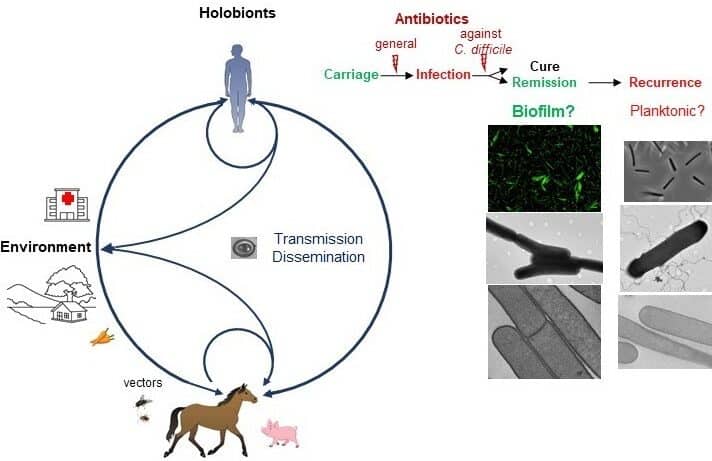
Nous étudions la formation et la structure des biofilms de Cd et leur rôle dans le cycle infectieux. Tout d’abord, dans une souche humaine modèle, le biofilm s’est avéré hétérogène avec des agrégats de cellules et une sous-expression des gènes de toxines (doi : 10.3389/fmicb.2018.02084). L’architecture du biofilm et la morphologie des cellules dans différentes conditions sont analysées, ainsi que l’induction du biofilm par des peptides antimicrobiens en collaboration avec l’équipe BaPS de Micalis et l’Institut Pasteur. Deuxièmement, en collaboration avec l’ANSES, des souches de Cd non domestiquées ont été collectées chez les équidés et caractérisées pour leur diversité moléculaire et leurs phénotypes, tandis que les animaux ont été diagnostiqués comme infectés ou porteurs. Dans le cadre d’un projet de doctorat, les biofilms des souches équines sont étudiés pour leur biomasse, leur architecture, leur anti-tolérance, leur production de toxines et de spores. En parallèle, avec le CNR Cd et les CHU, les génomes des souches équines et cliniques de la même région sont comparés pour surveiller la transmission intra- et inter-espèces. Cela permettra de comprendre la contribution des biofilms de Cd à la persistance, à la virulence et à la propagation, dans une perspective de santé unique.

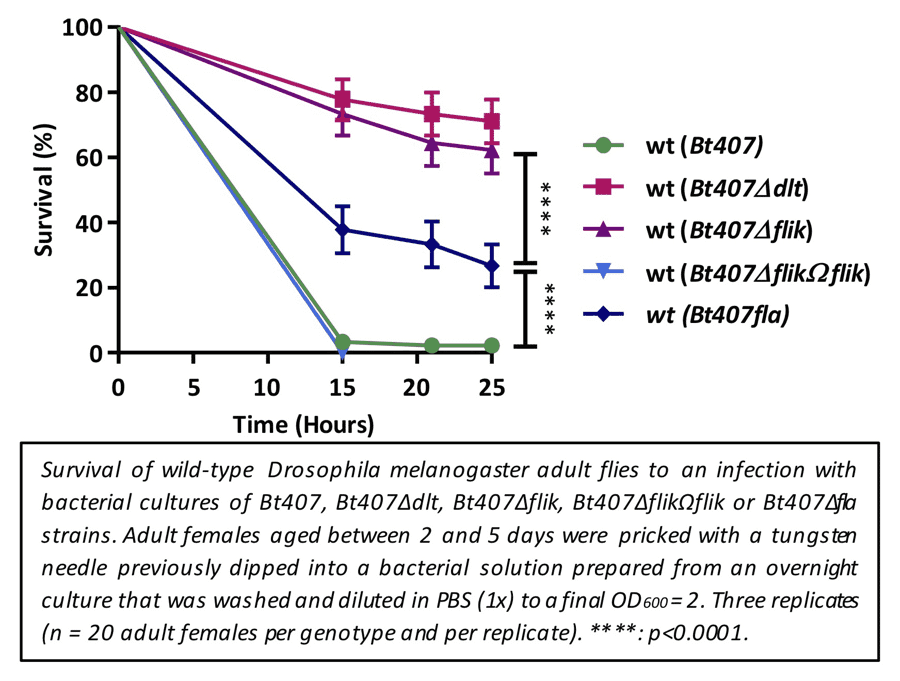
Les insectes possèdent des mécanismes efficaces qui leur permettent de détecter et de neutraliser les infections microbiennes. Nous nous intéressons à la caractérisation des mécanismes de résistance du Bt/Bc aux composants humoraux ou cellulaires de l’immunité innée de l’hôte (en particulier les peptides antimicrobiens) et aux réponses immunitaires et physiologiques de l’hôte activées au niveau de la barrière épithéliale chez G. mellonella et chez Drosophila melanogaster lors d’une infection (en collaboration avec l’Université Saint-Joseph au Liban). Nous avons découvert que, outre son rôle dans la résistance aux peptides antimicrobiens cationiques (CAMPs), l’opéron dltXABCD des bactéries Gram-positives, qui est requis pour l’incorporation d’esters de D-alanine dans les acides teichoïques associés à la paroi cellulaire, limite la libération de fragments de peptidoglycane immunostimulants et leur détection ultérieure par les récepteurs du système immunitaire inné de l’hôte (doi : 10.3389/fmicb.2020.611220). Nous avons également identifié un nouveau facteur bactérien (FliK) impliqué dans la résistance du Bt aux composants humoraux ou cellulaires de l’immunité innée de la Drosophile et dans la modulation de sa réponse immunitaire (doi : 10.3389/fmicb.2020.611220 et doi : 10.1016/j.resmic.2023.104089). Ces travaux pourraient conduire au développement de nouvelles stratégies de lutte contre les insectes visant à améliorer l’efficacité des agents de biocontrôle en réduisant la compétence immunitaire de l’hôte, ainsi qu’à la recherche de nouvelles cibles thérapeutiques comme alternative aux antibiotiques.
Le groupe B. cereus comprend des espèces étroitement apparentées qui produisent des facteurs de virulence, tels que des entérotoxines, des hémolysines, des phospholipases, des protéases et des adhésines (doi : 10.1128/microbiolspec.GPP3-0032-2018). Le régulateur de virulence principal PlcR, qui appartient à la famille des capteurs de quorum RRNPP, contrôle l’expression de la plupart de ces facteurs. Cependant, alors que certaines souches sont responsables de gastro-entérites d’origine alimentaire, il n’y a pas de corrélation entre la présence et le niveau d’expression des gènes de virulence et la pathogénicité des souches. La distinction des souches entéropathogènes est un défi en raison de l’absence de modèle animal disponible. Notre objectif est d’étudier la régulation de la production de facteurs de virulence chez les souches diarrhéiques par rapport aux souches environnementales dans des conditions similaires à celles que l’on trouve dans l’intestin. Une meilleure compréhension de la régulation de la pathogénicité de Bc permettra d’identifier les déterminants génétiques spécifiques qui différencient les souches entéropathogènes des autres.
Notre groupe a identifié un nouvel acteur de la virulence de Bt : le régulateur transcriptionnel VipR (doi : 10.1128/microbiolspec.GPP3-0032-2018). Ce régulateur est autorégulé et contrôle l’expression de la protéine insecticide exportée Vip3A, une toxine très active vis-à-vis de l’insecte Spodoptera frugiperda, un ravageur des cultures. En plus de Vip3A, VipR active plusieurs autres gènes au début de la phase stationnaire, y compris les gènes de l’amidase et de la toxine Cry. L’ensemble de ces résultats conduit à reconsidérer la régulation et le rôle des toxines Cry au cours de l’infection, que 40 ans d’études avaient définitivement classées comme des protéines spécifiquement produites au cours de la sporulation pour former des cristaux associés aux spores. Nous étudions actuellement le rôle de ce régulateur et des amidases, ainsi que les fonctions potentiellement inattendues des toxines insecticides exprimées précocement au cours du cycle infectieux de la bactérie dans un modèle d’insecte. Une partie de ce travail sera menée dans le cadre du LIA BIPI axé sur les interactions bactéries-plantes-insectes pour le biocontrôle des maladies.
 La plupart des souches émétiques de Bc hébergent un mégaplasmide appelé pCER270. Ce plasmide porte le locus ces impliqué dans la production de la toxine émétique, ainsi que de nombreux gènes de fonction inconnue ou codant pour des régulateurs. Nous avons montré que pCER270 est impliqué dans des phénotypes spécifiques aux souches émétiques, tels que la résistance élevée des spores à la chaleur ou la formation de biofilms atypiques, et nous émettons l’hypothèse que ces phénotypes spécifiques rendent les souches émétiques bien adaptées à l’alimentation humaine et contribuent donc à leur pathogénicité. Par conséquent, le transfert de pCER270 à des souches non pathogènes pourrait entraîner l’émergence de nouveaux pathogènes. Bien que pCER270 ne soit pas un plasmide conjugatif, nous avons réussi à le transférer à Bt et B. weihenstephanensis par un processus de conduction et avons constaté que le transfert du plasmide avait un impact sur le transcriptome du nouvel hôte (doi : 10.1016/j.resmic.2023.104074). Nous avons également constaté que pCER270 a un impact sur la sporulation et la formation de biofilms et nous avons identifié les régulateurs codés par pCER270 impliqués dans les phénotypes induits par pCER270. Nous déterminons actuellement l’effet de pCER270 sur l’aptitude de son hôte dans divers environnements.
La plupart des souches émétiques de Bc hébergent un mégaplasmide appelé pCER270. Ce plasmide porte le locus ces impliqué dans la production de la toxine émétique, ainsi que de nombreux gènes de fonction inconnue ou codant pour des régulateurs. Nous avons montré que pCER270 est impliqué dans des phénotypes spécifiques aux souches émétiques, tels que la résistance élevée des spores à la chaleur ou la formation de biofilms atypiques, et nous émettons l’hypothèse que ces phénotypes spécifiques rendent les souches émétiques bien adaptées à l’alimentation humaine et contribuent donc à leur pathogénicité. Par conséquent, le transfert de pCER270 à des souches non pathogènes pourrait entraîner l’émergence de nouveaux pathogènes. Bien que pCER270 ne soit pas un plasmide conjugatif, nous avons réussi à le transférer à Bt et B. weihenstephanensis par un processus de conduction et avons constaté que le transfert du plasmide avait un impact sur le transcriptome du nouvel hôte (doi : 10.1016/j.resmic.2023.104074). Nous avons également constaté que pCER270 a un impact sur la sporulation et la formation de biofilms et nous avons identifié les régulateurs codés par pCER270 impliqués dans les phénotypes induits par pCER270. Nous déterminons actuellement l’effet de pCER270 sur l’aptitude de son hôte dans divers environnements.
Certains insectes sont considérés comme une nouvelle source durable de protéines pour la consommation humaine et animale, parmi lesquels le ver de farine Tenebrio molitor et les larves de la mouche soldat noire, Hermetia illucens. Galleria mellonella sert de modèle pour étudier les interactions entre le microbiote, le pathogène et les réponses de l’hôte. Nos recherches portent sur deux aspects principaux : 1) la sécurité microbienne des aliments à base d’insectes et 2) le rôle du microbiote intestinal et de la qualité du substrat alimentaire dans la résistance des insectes aux agents pathogènes.

Afin de proposer une alternative aux pesticides chimiques, nous avons développé un nouveau type de souches biopesticides destinées à être utilisées contre les insectes ravageurs des cultures et les vecteurs de maladies (BIOSAFE, MOSKO, BT-VIP). Avec le soutien financier de la SATT Paris-Saclay et de l’INRAE, ces souches ont été développées en utilisant les résultats générés par nos travaux sur les régulateurs en phase stationnaire tels que CpcR et VipR, mentionnés ci-dessus, ainsi que des brevets (EP17305011 et PCT/EP2023/061003).
Bt est largement utilisé dans la protection des cultures en raison de sa production massive de protéines insecticides Cry, stockées dans de grands corps cristallins parasporaux. Nous avons utilisé les outils moléculaires de Bt pour créer une nouvelle plateforme de production de protéines hétérologues et de cristallisation in vivo. En utilisant la GFP comme preuve de concept pour notre plateforme, nous avons découvert que cette protéine était stockée dans des cristaux fluorescents et qu’elle représentait jusqu’à 60 % du contenu protéique total de la bactérie. Nous améliorons actuellement le processus pour le rendre utilisable avec une large gamme de protéines. Les résultats de ce projet sont multiples.La cristallisation in vivo facilitera la détermination de la structure 3D des protéines en évitant le processus long et fastidieux de cristallisation des protéines in vitro.
En outre, les protéines cristallisées sont stables et présentent un faible mouvement intramoléculaire, ce qui permet de produire des protéines instables ou cytotoxiques. Enfin, les cristaux peuvent être utilisés comme vecteurs de médicaments pour une libération lente des produits pharmaceutiques. Cette technologie est basée sur des résultats brevetés (PCT/EP2023/061003 et PCT/EP2023/061001).
Bien que la plupart des souches de Bc soient inoffensives et largement répandues dans l’environnement, un certain nombre de souches sont impliquées dans des intoxications alimentaires. En effet, Bc est l’un des micro-organismes les plus fréquemment isolés dans les épidémies d’origine alimentaire. Cependant, les marqueurs moléculaires ne sont disponibles que pour les souches émétiques produisant la toxine cereulide ou pour les souches portant le gène cytK1 codant pour la cytotoxine K1, qui représentent toutes deux moins d’un tiers des cas d’intoxication alimentaire. Tous les autres facteurs de virulence se retrouvent aussi bien dans les souches environnementales que dans les souches pathogènes. Par conséquent, presque tous les isolats provoquant des symptômes diarrhéiques ne peuvent pas être actuellement détectés ou identifiés. Nous émettons l’hypothèse que la quantité de toxines produites, plutôt que la présence d’un facteur de virulence spécifique, est l’un des facteurs critiques responsables de la pathogénicité des souches diarrhéiques.Nous sommes donc engagés dans la mise en place de tests immuno-enzymatiques afin de quantifier la production de facteurs de virulence par des souches suspectes de Bc.
Nous avons déjà développé avec succès deux tests ELISA, l’un dirigé contre la phospholipase sphingomyélinase, l’autre contre l’hémolysine HlyII.

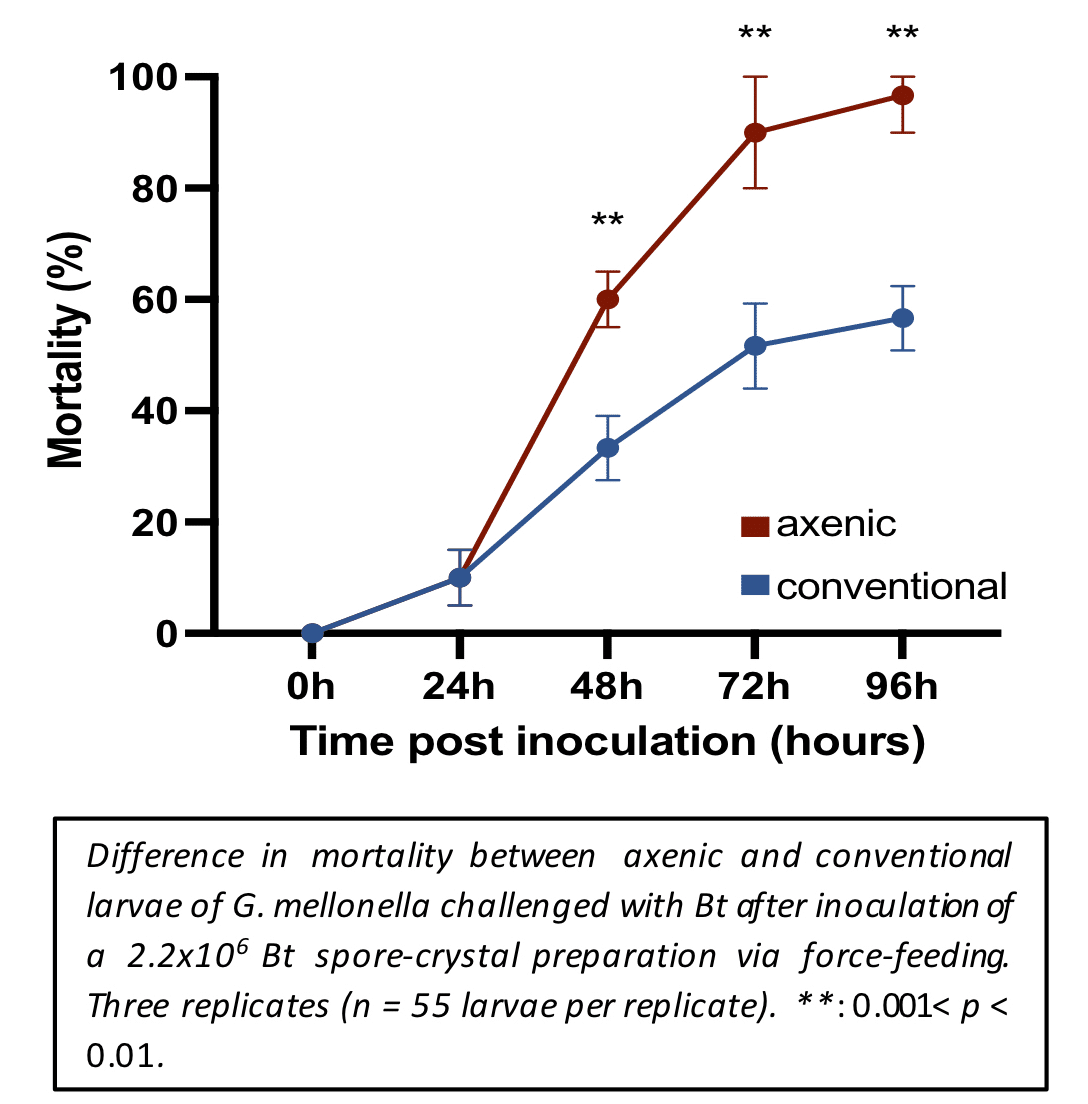
Les stades larvaires de l’insecte lépidoptère G. mellonella sont de plus en plus utilisés dans de nombreux laboratoires comme modèle d’infection pour les pathogènes bactériens et fongiques. Pouvant être utilisé pour des études précliniques, il répond au concept des 3R (réduire, remplacer, optimiser) afin de limiter l’utilisation de modèles mammifères, notamment pour le criblage de nouveaux antibiotiques mais aussi pour les études d’interaction entre le pathogène et l’hôte. Cet insecte est élevé dans notre insectarium depuis de nombreuses années et a été utilisé pour identifier des gènes impliqués dans la virulence, l’adaptation et la survie et pour visualiser leur expression in situ, en particulier pour Bc et Bt (voir résultats dans les Axes 1, 2 et 3). Des études hôte-pathogène-microbiote ont également été entreprises car nous sommes en mesure d’élever des larves axéniques et de réaliser l’infection par gavage oral. Nous produisons et vendons les derniers stades de G. mellonella à plusieurs laboratoires scientifiques en France. Nous avons mis en place des collaborations pour étudier notamment les interactions entre les bactéries et le tractus digestif en ce qui concerne le rôle du microbiote intestinal et de la matrice péritrophique, en utilisant des approches de dissection et d’histologie. Par exemple, en utilisant des larves axéniques, nous avons montré que le microbiote intestinal ne jouait aucun rôle dans la croissance de G. mellonella sur sa nourriture naturelle (cire d’abeille) et que la larve n’était pas capable d’assimiler le polyéthylène plastique (PE) comme cela a été montré en collaboration avec le Synchrotron en utilisant la microscopie infrarouge FTIR sur des sections de larves entières nourries avec du PE marqué (voir figure) (doi : 10.1021/acs.est.1c03417).




















